If the HBG are not prone to recruitment amongst themselves, nor are they likely to cross and backcross, where did the multitude of stories regarding reversion to GSF come from?
Sprk a scientific reply is - there is a whole lot we don't know. Bill C. gave you at least part of the answer - multi species , reciprocal cross differences , % of pure genetics , regional adaptation and more. I start my analysis with the following.
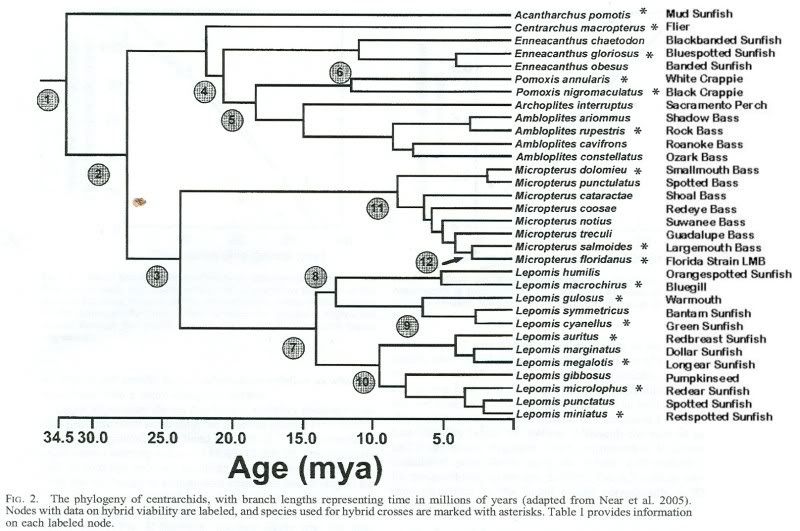
TEMPO OF HYBRID INVIABILITY IN CENTRARCHID FISHES
By DANIEL I. BOLNICK AND THOMAS J. NEAR
Received September 13, 2004. Accepted February 15, 2005.*
Abstract. Hybrid viability decreases with divergence time, a pattern consistent with a so-called speciation clock.
However, the actual rate at which this clock ticks is poorly known. Most speciation-clock studies have used genetic
divergence as a proxy for time, adopting a molecular clock and often far-distant calibration points to convert genetic
distances into age. Because molecular clock assumptions are violated for most genetic datasets and distant calibrations
are of questionable utility, the actual rate at which reproductive isolation evolves may be substantially different than
current estimates suggest. We provide a robust measure of the tempo at which hybrid viability declines with divergence
time in a clade of freshwater fishes (Centrarchidae). This incompatibility clock is distinct from a speciation clock
because speciation events in centrarchids appear to be driven largely by prezygotic isolation. Our analyses used
divergence times estimated with penalized likelihood applied to a phylogeny derived from seven gene regions and
calibrated with six centrarchid fossils. We found that hybrid embryo viability declined at mean rate of 3.13% per
million years, slower than in most other taxa investigated to date. Despite measurement error in both molecular
estimated ages and hatching success of hybrid crosses, divergence time explained between 73% and 90% of the
variation in hybrid viability among nodes. This high correlation is consistent with the gradual accumulation of many
genetic incompatibilities of small effect. Hybrid viability declined with the square of time, consistent with an increasing
rate of accumulation of incompatibilities between divergent genomes (the snowball effect). However, the quadratic
slope is due to a lag phase resulting from heterosis among young species pairs, a phenomenon rarely considered in
predictions of hybrid fitness. Finally, we found that reciprocal crosses often show asymmetrical hybrid viabilities.
We discuss several alternative explanations for this result including possible deleterious cytonuclear interactions.
Speciation-clock studies have been a small cottage industry recently, but there are still novel insights to be gained
from analyses of more taxonomic groups. However, between-group comparisons require more careful molecular-clock
calibration than has been the norm.
we have not been able
to find any documentation of F1 fertility for taxa more than
14.64 million years apart.
In contrast, the minimum age for total inviability
in centrarchids is 24.81 million years (node 3, Fig. 2), though
this is only for one direction of a reciprocal cross. The other
direction of this cross (L. microlophus 3 M. salmoides) yielded
43% viability. The only crosses with total inviability in
both directions are M. salmoides 3 (Ambloplites rupestrus,
Pomoxis annularis, or Pomoxis nigromaculatus) at 28.94 million
years, while 10 other crosses of that age have some
viability in one or both reciprocal directions (see online Appendix).
Centrarchids also retain nonzero viability and heterosis
for much longer than most other taxa.
We speculate that the slow evolution of hybrid inviability in
centrarchids reflects a lack of distinctive large hemizygous
sex chromosomes, preventing the more rapid accumulation
of inviabilities via Haldane�s rule (Turelli and Begun 1997).
The asymmetrical viability of reciprocal F1 hybrids argues
for a major role of interactions between haploid (sex or mitochondrial
loci) and diploid genes, or cytonuclear interactions
in postzygotic isolation. Previous studies of allozyme
gene expression ontogeny in hybrid centrarchids lend support
to the latter effect (Whitt et al. 1977). Our results also suggest
that theory needs to assess the interaction between heterosis
and genetic incompatibilities before we can judge whether a
lag phase (Mendelson et al. 2004) in the evolution of hybrid
inviability can be used to support the snowball effect (Orr
and Turelli 2001). Finally, we argue that the evolution of
hybrid inviability plays little role in driving speciation in
centrarchids. Nonetheless, inviability may still play an important
role in buttressing species against subsequent introgression.
This is because other reproductive isolating mechanisms
can break down under habitat disturbance or environmental
change, whereas intrinsic genetic incompatibilities
guarantee that diverged lineages remain distinct.
We do not currently have enough information to distinguish
between sex chromosome, mitochondrial, or cytoplasmic
effects. However, the lack of distinctive sex chromosomes
(Roberts 1964; but see Becak et al. 1966) suggests
that the hemizygous nuclear region is likely to be small (possibly
even a single locus) and so may not contribute strongly
to inviability (Turelli and Begun 1997). One puzzling pattern to emerge from our data lends some credence to a role for
cytonuclear interactions: using maximum body size as an
index (Page and Burr 1991), the larger species tends to be
the more successful maternal parent (Table 3). Of the 18
species pairs with reciprocal cross data and nonzero viability,
one pair had equal body size and nearly symmetrical crossing
success. Focusing on the remaining 17 species pairs (admittedly
not phylogenetically independent; Table 3), the larger
parent was more successful in 13 crosses and less successful
in four crosses (x12 5 4.765, P 5 0.029). We speculate that
there is greater disruption of paternal allele expression when
the paternal allele is from a smaller species, placed in an egg
with cytoplasmic factors encoded by a larger maternal species.
However, the cytoplasmic effect cannot be attributed to
differences in egg size, as egg size is not correlated with
body size (D. I. Bolnick, unpubl. data) and egg size differences
are not associated with inviability (Merriner 1971b).
We are working on expanding our dataset to include more
reciprocal crosses to test this pattern more rigorously.





.gif)







