Old , very old data and a lot of water (studies ) under the bridge since this article. Some right , some wrong , some out of context. Where to start.
With the concept of outbreeding depression which all those crosses exhibit and the article did not touch (or intentionally left out ). I guaranty Childers was well aware of this as he is/was a preeminent geneticist in lepomis. Gen one may (or may not depending on perspective ) be superior to the parentals. Gen 2 is absolutely not superior to parentals. Gen 3 worse - you get the pic.
Hybrid lepomis can not successfully be the forage base for LMB except for a couple of non existent crosses.
While it is possible and does happen (more so in lepomis) most times reciprocal crosses (GSFm X RESf and RESm X GSFf ) have similar fecundity. One thought on the wide variance of results with GSF crosses is the difficulty of finding pure GSF stock. GSF are the most likely lepomis to cross with other species.
Here are the sex ratios (percent male) that Childers got with various hybrid crosses. The male parent is listed first. There is some variation in the numbers from other studies
Redear X Bluegill 97 (3)
Bluegill X Redear 97
Redear X Green 69
Green X Redear 48
Bluegill X Green 97
Green X Bluegill 68 (2)
Redear X Warmouth 55
Bluegill X Warmouth 69 (2)
Green X Warmouth 16
Warmouth X Green 84
Note which species has the consistentaly skewed reciprocal cross data in this list.
HYBRIDIZATION OF FISHES IN NORTH AMERICA
(FAMILY CENTRARCHIDAE)
by
W.F. CHILDERS
Illinois Natural History Survey
Urbana, Illinois
U.S.A.
Sex Ratios of F1 hybrids
Sexually mature F1 hybrids were collected from each population and sexed. Of the 10 kinds of viable F1 hybrids, seven were predominately males (RB, BR, and BG were 97 percent males; WG were 84 percent males; and RG, GB, and BW were approximately 70 percent males), two were approximately 50 percent males (GR and RW), and one was predominately female (GW was 16 percent males). Ricker (1948) determined the sex of 428 BR F1 hybrids in Indiana and found them to be 97.7 percent males.
Sex determination in sunfishes is very poorly understood. Bluegills, green sunfish, and their F1 hybrids apparently have 24 pairs of chromosomes, and the sex chromosomes are indistinguishable from the autosomes (Bright 1937). Bright also reported that the chromosomes are so similar in shape and size that he was unable to detect specific differences. Roberts (1964) found that red-ear, bluegill, and warmouth sunfishes each have 24 pairs of chromosomes; green sunfish from North Carolina had 24 pairs; but green sunfish from West Virginia had only 23 pairs.
The unbalanced phenotypic tertiary sex ratios of the F1 hybrid sunfish could result from unbalanced primary genetic sex ratios, specific differences in the strength of sex-determining factors, an overriding of the genetic sex by environmental factors, or differential mortality of the sexes.
Since the WG F1 hybrids were 84 percent males and the reciprocal cross hybrids were 16 percent males, it is possible that the strength of sex-determining factors of warmouths are 5.25 times more powerful than those of green sunfish. Specific differences in the strength of sex-determining factors cannot alone explain the sex ratios of the remaining eight kinds of viable hybrids, since none of these were predominately females.
RB and BG F1 hybrids were both 97 percent males. If differential mortality were the cause of these unbalanced sex ratios, much of the mortality would have had to occur after the swim-up fry stages, since in the stripping experiments total mortality between fertilization and the swim-up fry stages was only 14 percent for the RB and 27 percent for the BG F1 hybrids.
It is not known which sex is the heterogametic condition for the sex chromosomes of the four experimental species; however, Haldane (1922) formulated a rule which furnishes a clue: �When in the F1 offspring of a cross between two animal species or races, one sex is absent, rare, or sterile, that sex is always the heterozygous sex.� Using Haldane's rule, Krumholz (1950), in a study concerning BR F1 hybrids, pointed out that the males of both bluegills and red-ear sunfish are probably homozygametic for sex and the females heterozygametic. The application of Haldane's rule to all possible F1 hybrids produced from red-ear sunfish, bluegills, and green sunfish indicates that the female is the heterozygametic sex in these three species. Hybridization of male warmouths with females of the three Lepomis species resulted in partial or complete lethals, suggesting that in the warmouth the male is the heterogametic sex.
Some of this has since been proven wrong.
NorthA merican Journal of Fisherie Msnagemen6t: 156-167, 1986
� Copyrighbt y the AmericanF isheriesS ociety1 986
Evaluation of Male Bluegill X Female Green Sunfish Hybrids for
Stocking Mississippi Farm Ponds
MARTIN W. BRUNSON and H. RANDALL ROBINETTE
The use of hybrid sunfishes in ponds has been
suggesteda s an attractive alternative to the more
traditional stocking policies involving the bluegill
(Lepomism acrochirus)o r redears unfish( Lepomis
microlophus), or both, in combination with the
largemouth bass (Micropterus salmoides) (Lewis
and Heidinger 1978a).
Ellison and Heidinger
(1978) surveyed 30 privately owned hybrid
sunfish ponds in southern Illinois and determined
that the presence of largemouth bass and supplemental
feeding were the two most important variables
related to growth of hybrids.
Growth of redear
sunfish x green sunfish and bluegill x green
sunfish (BG x GS) hybrids exceeded the average
growth for bluegill in Illinois (Lopinot 1972). The
BG x GS hybrids reached an acceptable catch size
in a shorter time than channel catfish (Ictalurus
punctatus), and they were easier to catch where
both were stocked for recreational fishing.
Several hybrid combinations have been recognized
as having potential in pond management
(Lewis and Heidinger 1978b), but the BG x GS
hybrid appears to have the most attractive combination
of desirable attributes. Like other hybrids,
the BG x GS hybrid exhibits rapid growth
(Childers 1967; Ellison and Heidinger 1978) and
probable hybrid vigor (Brunson and Robinette
1985), produces mostly males (Childers and Bennett
1961; Laarman 1973; Brunson 1983), and is
highly vulnerable to capture by hook and line
(Henderson and Whiteside 1976; Crandall and
Durocher 1980; Brunson 1983). Only the BG x
GS hybrid, however, is well suited to artificial
feeding (Lewis and Heidinger 1978a) and frequents
shallow water areas, where it may be more
accessiblet o the pond angler than some other hybrids.
The BG x GS hybrid is probably the most
common commercially produced hybrid sunfish
in the southeastern United States.
From an angler's viewpoint, increased growth is
highly desirable.H owever, perhapsm ore important
to the angler is the presumed high catchability of
hybrid sunfish. This vulnerability to hook-andline
capture has been alluded to by many authors
but relatively few experimental data exist to substantiate
such statements. Childers (1967) cited
one instance where a population of more than
10,000 BG x GS hybrids was decimated by anglers
during the first week of fishing. Childers and
Bennett (1967) reported hook-and-line yields in a
0.4-hectare pond stocked with redear x green sunfish
hybrids and largemouth bass that surpassed
those of comparably fished bluegill ponds. They
concluded that hybrids were more aggressive, less
wary, and less able to learn how to avoid being
caught than their parent species. Henderson and
Whiteside (1976) confirmed the vulnerability of
hybrids to angling, and Ellison and Heidinger
(1978) reported that hybrid sunfish were much
easier to catch than channel catfish when both were
stocked for recreational fishing. They also confirmed
the aggressivenesos f hybrid sunfisha s reported
by swimmers who were nipped by these
fish. Crandall and Durocher (1980) reported that
catchability of the BG x GS hybrid was significantly
higher than that ofbluegill x redear sunfish
or green x redear sunfish hybrids.
Other than theses tudies,t he literature on hybrid
sunfishes is the result of laboratory or controlled,
short-term pond studies with relatively narrow objectives.
The total catch of
146 hybrids during the 2-h period was 21% of the
population.
These high catch rates,
especially at Britt Pond, once again confirm the
aggressivenessa nd vulnerability of the BG x GS
hybrid, and point to the importance of strict control
of harvest in hybrid ponds. Especially significant
is the effect that poachers can have on a
hybrid population.
Though hybrid sunfish are not a panacea
for all farm pond management problems, they can
be used under certain conditions to produce desirable
populations of rapidly growing fish. With
a rapid growth rate, low population fecundity, and
high catchability, these fish can be used to create
high quality bream fishing when properly managed
TEMPO OF HYBRID INVIABILITY IN CENTRARCHID FISHES
(TELEOSTEI: CENTRARCHIDAE)
DANIEL I. BOLNICK1,2 AND THOMAS J. NEAR3,4
Asymmetries in F1 Hybrid Viability
Centrarchid hybrid viability differs between reciprocal
crosses of the same pair of species (F1 asymmetry). Of 18
species pairs for which reciprocal crosses have been done
(and viability is nonzero), 17 had significantly different viabilities
depending on which species was the female (or male)
parent. The relative strength of this asymmetry increased linearly
with time (Fig. 4), because the absolute difference in
viabilities was fairly constant and represented an increasing
proportion of the overall viability as the latter measure declined.
Asymmetrical F1 viabilities may also result from deleterious
interactions between the maternally provided oocyte
cytoplasm and the hybrid�s nuclear genes. Centrarchid hybrids
show aberrant timing of allozyme gene expression during
early development, even when the parental species have
identical onset of gene expression (Phillip et al. 1983). These
results suggest that centrarchid species have diverged in their
gene regulation mechanisms even while expression location
and timing remained similar. In many cases, hybrids expressed
maternal alleles at the normal time, but paternally
derived alleles were delayed, premature, or failed to be expressed
at all (Phillip et al. 1983). Less viable hybrids in a
reciprocal cross are generally the ones with greater paternal
allele misexpression. Whitt et al. (1977) suggested that the
greater effect on paternal alleles is evidence for cytoplasmicnuclear
interactions, hypothesizing that maternally encoded
regulatory signals are misinterpreted by the paternal allele.
If one species� gene expression is more sensitive to changes
in transcription factors, asymmetries will result.
One puzzling pattern
to emerge from our data lends some credence to a role for
cytonuclear interactions: using maximum body size as an
index (Page and Burr 1991), the larger species tends to be
the more successful maternal parent (Table 3). Of the 18
species pairs with reciprocal cross data and nonzero viability,
one pair had equal body size and nearly symmetrical crossing
success. Focusing on the remaining 17 species pairs (admittedly
not phylogenetically independent; Table 3), the larger
parent was more successful in 13 crosses and less successful
in four crosses (x12 5 4.765, P 5 0.029). We speculate that
there is greater disruption of paternal allele expression when
the paternal allele is from a smaller species, placed in an egg
with cytoplasmic factors encoded by a larger maternal species.
However, the cytoplasmic effect cannot be attributed to
differences in egg size, as egg size is not correlated with
body size (D. I. Bolnick, unpubl. data) and egg size differences
are not associated with inviability (Merriner 1971b).
We are working on expanding our dataset to include more
reciprocal crosses to test this pattern more rigorously
HAVING POSTED THIS ON CENTRAS NOTE THAT RES X BG SEEM TO BE THE EXCEPTION FOR SEVERAL REASONS INCLUDING NEAR SIMILAR SIZE OF THE TWO SPECIES AND DO CROSS WELL IN BOTH DIRECTIONS.
TEMPO OF HYBRID INVIABILITY IN CENTRARCHID FISHES
(TELEOSTEI: CENTRARCHIDAE)
DANIEL I. BOLNICK AND THOMAS J. NEAR
Centrarchid hybrids
show aberrant timing of allozyme gene expression during
early development, even when the parental species have
identical onset of gene expression (Phillip et al. 1983). These
results suggest that centrarchid species have diverged in their
gene regulation mechanisms even while expression location
and timing remained similar. In many cases, hybrids expressed
maternal alleles at the normal time, but paternally
derived alleles were delayed, premature, or failed to be expressed
at all (Phillip et al. 1983). Less viable hybrids in a
reciprocal cross are generally the ones with greater paternal
allele misexpression. Whitt et al. (1977) suggested that the
greater effect on paternal alleles is evidence for cytoplasmicnuclear
interactions, hypothesizing that maternally encoded
regulatory signals are misinterpreted by the paternal allele.
If one species� gene expression is more sensitive to changes
in transcription factors, asymmetries will result.
the larger species tends to be
the more successful maternal parent (Table 3). Of the 18
species pairs with reciprocal cross data and nonzero viability,
one pair had equal body size and nearly symmetrical crossing
success. Focusing on the remaining 17 species pairs (admittedly
not phylogenetically independent; Table 3), the larger
parent was more successful in 13 crosses and less successful
in four crosses ( 5 4.765, P 5 0.029). We speculate that 2 x1
there is greater disruption of paternal allele expression when
the paternal allele is from a smaller species, placed in an egg
with cytoplasmic factors encoded by a larger maternal species.
However look at GSF X BG below where the better dam is not the larger species ( one of only a few).
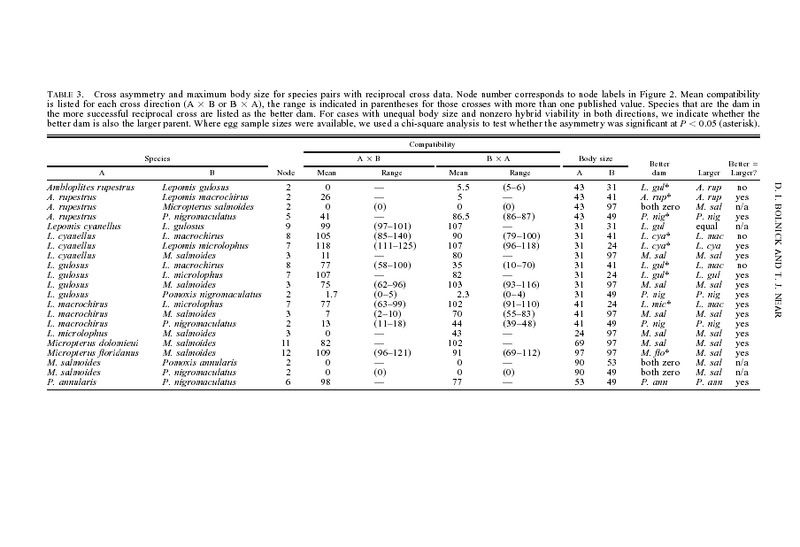
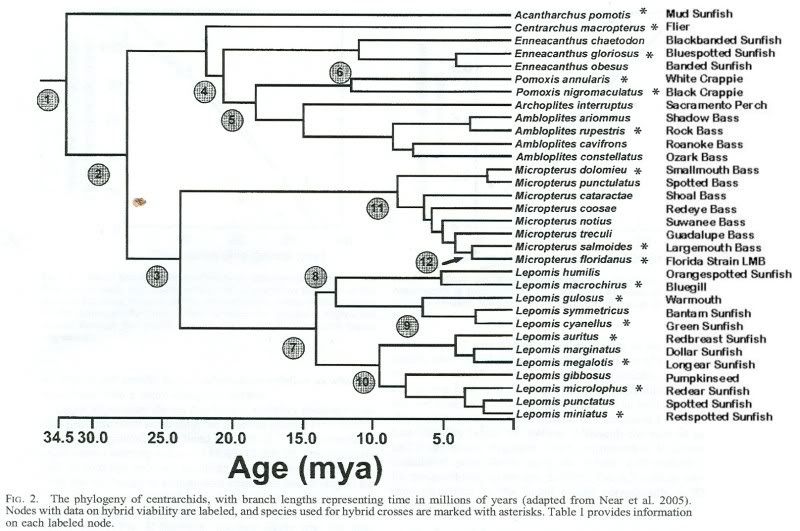
TEMPO OF HYBRID INVIABILITY IN CENTRARCHID FISHES by DANIEL I. BOLNICK AND THOMAS J. NEAR :
" The only crosses with total inviability in
both directions are M. salmoides 3 (Ambloplites rupestrus,
Pomoxis annularis, or Pomoxis nigromaculatus) at 28.94 million
years, while 10 other crosses of that age have some
viability in one or both reciprocal directions (see online Appendix).
Centrarchids also retain nonzero viability and heterosis
for much longer than most other taxa."







.gif)


![[Linked Image from i1261.photobucket.com]](http://i1261.photobucket.com/albums/ii582/teehjaeh57/I_subscribe_zps69249d43.gif)

![[Linked Image]](http://forums.pondboss.com/ubbthreads.php?ubb=download&Number=11852&filename=I_subscribe_zps2qlyoyn2.gif)







