Walt do you have those studies ? I have seen shad added to many lakes/ponds with all different outcomes. I have read many studies on the subject both as to GShad and Tshad. Generalizations are suspect. I have added below some text which should show the uncertainty and differences. From these I gather that GShad are a problem unless you
want and have big LMB as noted above. TShad are not the same as they often don't compete significantly with BG and don't suppress the recruitment of LMB and BG like GShad. Even in the GShad studies it is not so much the competition for food as it is reduced predation on BG and other factors. The last study below is an examination of some 60 prior studies on GShad and TShad as forage .
The only 2 methods I know of for trophy BG ponds are LMB crowded ponds or all male BG ponds. Trophy HBG ponds are a combo of those 2.
Direct and indirect effects of gizzard shad on bluegill growth and population size structure
"These results demonstrate that the presence of gizzard shad is associated with reduced bluegill growth rates and adult size structure and
that mechanisms other than direct competition for food resources may be responsible"
Effects of Waterbody Type and Management Actions on Bluegill Growth Rates
"
Managers should recognize basic differences in growth patterns between water body types when determining which management actions should be implemented or when evaluating their success."
The Influence of Gizzard Shad (Dorosoma clpedianum) on Plankton Community Dynamics in Earthen Ponds
Date: 1995
Abstract: The impacts of gizzard shad on plankton community interrelationships were evaluated in six, 2 to 5 ha ponds over a 3-year period. All ponds were stocked with largemouth bass (Micropterus salmoides) and sunfish (Lepomis spp.). Three of the six ponds were stocked with gizzard shad at random while the other three served as controls. The zooplankton and phytoplankton were sampled and analyzed three times during 1992 (before shad introduction) and twice a month during the growing seasons of 1993 and 1994. Water quality variables were measured once in 1992 and every other month during the growing seasons of 1993 and 1994. Phytoplankton primary productivity was measured three times in 1992 before the introduction of shad and once a month during the growing seasons of 1993 and 1994. Zooplankton density, biomass and size were affected by gizzard shad during summer, 1993 and spring and fall of 1994. Statistical differences between treatments occurred in spring, 1994 (p<0.13) with higher density, higher biomass, and smaller organisms in the shad treatment. Phytoplankton density, diversity and primary productivity were all higher in the shad treatments (p<0.13) during fall, 1994. Shad influenced the plankton community in a web-like fashion directly impacting zooplankton and phytoplankton simultaneously and indirectly affecting phytoplankton by altering the zooplankton. Shad shifted phytoplankton size structure by first reducing large phytoplankton (>70 mm) in the shad treatment immediately after shad introduction in 1993 and enhancing small phytoplankton (C28 mm) a year after shad introduction (1994). Through this size structuring, shad increased the photosynthetic efficiency of the phytoplankton community to twice that of ponds without shad by summer, 1994 (p =0.067) and increased primary productivity to 1.5 times that of ponds without shad by fall, 1994 (p =0.098).
Relationships between trophic state and gizzard shad, bluegill sunfish, and largemouth bass populations in three Alabama impoundments
Date: 1989
Abstract: Three Alabama impoundments were studied to analyze the relationship between fertility levels and the population and community structures of several species of fish. Chlorophyll a concentrations were used to classify lake trophic status. Fish communities and populations were similar in structure between the two eutrophic systems and differed from the oligotrophic system. Eutrophic systems were shown to have a higher relative abundance of prey species, in particular gizzard shad. Overall production was lower in the oligotrophic systems. Community structure in the eutrophic systems reflected a greater influence of gizzard shad. Bluegill abundance doubled while gizzard shad numbers increased twentyfive-fold. Largemouth bass increased eight times in number. Prey populations were overcrowded, characterized by slow growth, poor condition, and reduced spawning, due to the high relative abundance of gizzard shad. Predator populations showed low recruitment and poor condition in smaller fish due to lack of suitable size prey.The relative abundance of predator species increased in the oligotrophic system. Bluegill were in better condition and had better distribution across size classes. Young of year and age one gizzard shad were absent from sampling throughout the year.
Title: The effects of threadfin shad as a forage species for largemouth bass in combination with bluegill, redear, and other forage species
Date: 1976
Abstract: Threadfin shad, Dorosoma petenense (Gunther), were stocked in ponds with an established population of bass, bluegill, and redear and in three ponds containing bass, golden shiners, and fathead minnows. Comparisons of fish populations were made before and after the first successful spawn of shad. The production of bass with bluegill and redear plus shad was essentially the same as with golden shiners and fathead minnows--59.2 pounds per acre and 58.7 pounds per acre, respectively. A significant increase in the growth and condition of bass was seen in the pond stocked with bluegill and redear after the first shad spawn while no change was seen in bass survival.
Bluegill and redear recruitment increased significantly after the shad population became established. This apparently was caused by a decreased rate of predation by bass. As a result, the catch per unit of effort of bluegill and redear decreased significantly after the introduction of shad.
Bass were in better condition after shad stocking while no change in the condition of bluegill and redear could be detected. In ponds stocked with forage minnows, fathead minnows disappeared the second summer after stocking. An F/C ratio (Forage/Carnivorous) of 2.7 and an A value for bass (percentage of bass over 10 inches in length) of 93.4 was computed for the pond, indicating a bass-crowded condition due to heavy predation on the forage species.From these data, the stocking of shad as additional forage in bass-bluegill-redear ponds cannot be recommended. The stocking of shad with bass and other forage species appears to offer some advantages to bass fishermen if the production of forage species can be increased.
Stocking Threadfin Shad: Consequences for Young-of-Year Fishes
DENNIS R. DEVRIES, ROY A. STEIN, and JEFFREY G. MINER
Ohio Cooperative Fish and Wildlife Research Unit,3 and Department of Zoology The Ohio State University, Columbus, Ohio 43210, USA
GARY G. MITTELBACH
Kellogg Biological Station, Michigan State University, Hickory Corners, Michigan 49060, USA
Abstract
Threadfin shad Dorosoma petenense are commonly introduced into reservoirs to supplement prey available to piscivorous fishes. To determine how early life stages of threadfin shad and their potential competitors and predators interact, we introduced this species into two Ohio lakes�Clark and Stonelick�and evaluated how its young of year influenced young-of-year bluegills Lepomis macrochirus and largemouth bass Micropterus salmoides. After adults were stocked in April, peak abundance of young-of-year threadfin shad occurred in August in both lakes.
Bluegills generally spawned earlier than threadfin shad, which apparently reduced competition between young of these species. In Clark Lake, young-of-year threadfin shad did not reduce zooplankton populations, but in Stonelick Lake, peak abundance of young-of-year threadfin shad was followed by a precipitous decline in zooplankton. Data on cladoceran birth rates indicated this decline was due to increased predation by threadfin shad. Survival of bluegills to a size at which they move into the littoral zone also declined in Stonelick Lake, perhaps because of the virtual elimination of zooplankton. Limited survival of bluegills in turn contributed to reduced growth of young-of-year largemouth bass dependent on them as prey. Given that zooplankton declined in one but not the other lake, interactions among young-of-year fishes due to annually introduced threadfin shad will likely vary among systems and years. Nonetheless, introduced threadfin shad could, in some systems in some years, negatively affect growth and recruitment of the very species they were meant to enhance.
Young-of-year threadfin shad, though their
abundance peaked in late summer, co-occurred at
low densities with limnetic young-of-year bluegills
during May through September in Clark Lake.
When both species co-occurred in the limnetic
zone, they ate only limnetic zooplankton; however,
diet overlap values were typically <0.50.
Once blue gills moved inshore, the potential for
competition with threadfin shad declined greatly.Diets of bluegills collected from the littoral zone
did not change between 1987 and 1988 in Clark
Lake; all prey types (littoral, limnetic, and cyclopoid
copepods) were eaten. Though collected inshore,
bluegills apparently moved far enough offshore
to consume some limnetic prey, but they ate
enough littoral prey to reduce overlap with threadfin
shad, which continued to feed entirely on limnetic
prey.
Consequently,
although competition between young-of-year
bluegills and young-of-year threadfin shad may
occur (particularly during August in Stonelick
Lake), the outcome of such interactions in a lake
ultimately depends on abundance and species
composition of the zooplankton community and
on the relative spawning times of the predator
fishes. As demonstrated by our results, these factors
vary among lakes and years, making generalization
difficult.Thus, interactions between limnetic young-ofyear
threadfin shad and bluegills may have a pronounced
negative effect on young-of-year largemouth
bass growth if they lead to reduced survival
of young-of-year bluegills in the limnetic zone and
then to reduced recruitment of bluegills to the littoral
zone. Additionally, slower growth may reduce
overwinter survival of young-of-year largemouth
bass if overwinter survival depends on body
size and fat reserves (Adams et al. 1982a, 1982b;
reviewed in Adams and DeAngelis 1987). As a
consequence, the very management practice intended
to enhance the fishery for adult piscivores
may reduce survival of the target species. Although
these negative effects are not direct (unlike
the positive effects of increased prey availability),
they could have substantial consequences over
several years of such management manipulation
Manipulating Shad to Enhance Sport Fisheries
in North America: An Assessment
DENNIS R. DEVRIES' AND ROY A. STEIN
Manipulating forage fish populations to enhance sport fisheries is a common management
practice. Here we review the literature dealing with manipulations of gizzard shad Dorosoma
cepedianum and threadfin shad D. petenense to assess whether or not this practice has been
successful. Shad introduction has tended to enhance predators, such as white crappie Pomoxis
annularis, black crappie P. nigromaculatus, and largemouth bass Micropterus salmoides, and negatively
affect presumed competitors, such as bluegill Lepomis macrochirus. However, responses
have not been consistent within a species: some studies document negative responses of predators
or positive responses of competitors to shad introduction. Depending on the study, target species
have experienced negative, neutral, and positive effects due to shad removal, making generalizations
impossible.
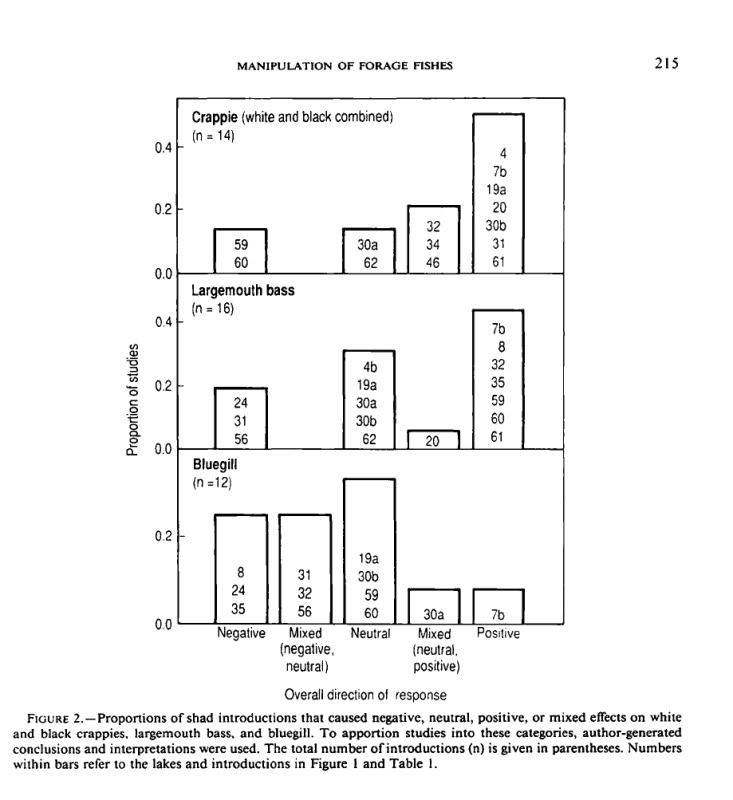
We were not able to generalize about how shad
influence sport fishes. Although bluegill appeared
to be more negatively affected by shad than crappies,
the response of largemouth bass being intermediate
between the two, we cannot draw definite
conclusions because the entire range of results occurred
for each target species.
The results presented here indicate that several
aspects of whole systems must be quantified if we
are to assess how forage-fish manipulations affect
a fish community. Based on our review of manipulations
involving gizzard and threadfin shad, potential
competition and predation, spatial refuges,
indirect effects through common predators and
prey, and the influence of ontogenetic shifts in
habitat and diet of the target and introduced species
are all critical to being able to predict the influence
of a forage-fish manipulation on a target species.





.gif)









